通过Atherton–Todd反应及衍生方法,研究构建了含44种BiPs与3种线性LiPs的化合物库(图1a-c),系统考察尾部链长(C14–C30)、分支位置、N-甲基化、极性头部及磷功能团(磷酰胺/膦酸酯/磷酸酯)对体内FLuc mRNA递送的影响;发现C20饱和支链尾部且分支点靠近磷官能团的磷酰胺脂质表现最优,其中BiP-20在CD1小鼠肝脏中产生最高生物发光信号(图1d),其LNP表观pKa为6.56(图1e),处于理想递送范围,且经DNA条形码配方优化后确认DOPE辅助脂质与常规摩尔比(50%离子化脂质/38.5%胆固醇/10% DOPE/1.5% PEG-脂质)适用于后续研究。
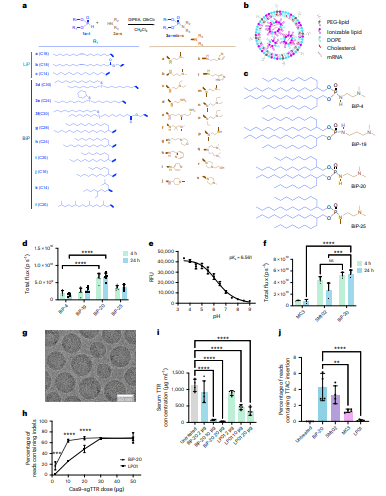
图 1. LiP和BiP脂质的化学合成、LNP筛选以及 CRISPR–Cas9 编辑系统的递送。
在CD1小鼠中,BiP-20 LNPs静脉注射后的荧光素酶表达较MC3 LNPs提高5.9倍,与SM102水平相当(图1f);机制研究表明其肝脏摄取高度依赖ApoE结合(ApoE⁻/⁻小鼠信号几乎完全消失)及LDL-R介导的内吞(LDL-R⁻/⁻小鼠信号显著降低),提示该递送系统通过经典脂蛋白受体通路实现肝细胞靶向。
在TTR基因编辑中,BiP-20 LNPs共包载Cas9 mRNA与sgTTR后呈现典型球形单层结构(图1g),在2 μg低剂量下即可实现约20%编辑,10 μg时达~64%,而LP01需30 μg方可比拟(图1h),血清TTR蛋白敲低与基因组编辑数据一致(图1i);在1 μg极低剂量下,BiP-20仍可产生约50%血清TTR降低与40% indel,显著优于MC3、SM102和LP01。在PCSK9 prime editing中(图1j),BiP-20在50 μg剂量下实现4.30%的TTAC插入率,显著优于SM102(3.33%)、MC3(1.33%)和LP01(0.1225%),证明其对复杂RNA共递送系统的高效性。
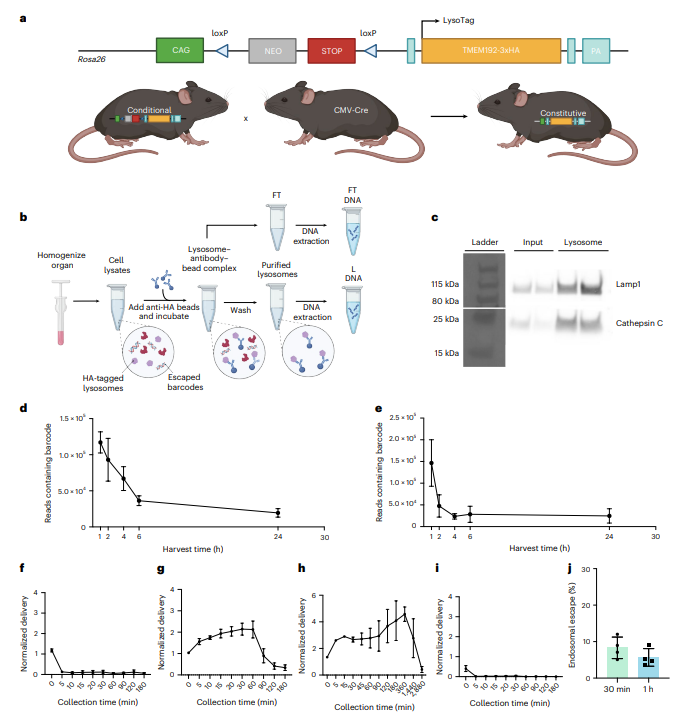
研究将floxed TMEM192-3×HA小鼠与CMV-Cre杂交获得组成性LysoTag小鼠(图2a),通过抗HA磁珠免疫沉淀从肝脏匀浆中分离纯化溶酶体,经Western blot验证Lamp1与Cathepsin C显著富集(图2c);全肝条形码可持续检测至24小时,但溶酶体内条形码在2小时内迅速衰减(图2d,e),提示早期即发生逃逸或降解。血液动力学显示BiP-20与裸条形码均迅速清除(图2f,i),而MC3和SM102分别稳定1小时和24小时(图2g,h),DiD标记证实BiP-20在5分钟内即被肝脏快速摄取。LysoBC定量显示,BiP-20 LNPs在给药后30分钟的内体逃逸效率约为8%,1小时降至约6%(图2j),流穿液经胞质标志物验证主要代表胞质内容。
图 2. 用于脂质纳米颗粒的体内溶酶体逃逸定量检测方法的发展。
在50 μg高剂量下,BiP-20处理小鼠的血清ALT与AST水平与未处理对照相当,无肝毒性;炎症因子检测中仅SM102显著诱导IL-6升高,BiP-20、LP01和MC3无显著变化,且各组TNF水平均未改变,表明BiP-20具有优异的体内耐受性。
蛋白质组学鉴定2,395种蛋白,发现BiP-20处理组有191种差异表达蛋白(DEPs),显著多于SM102的96种(图3a,b),且直接比较中BiP-20特异性下调7种蛋白(图3c);STRING分析显示BiP-20调控蛋白形成内体加工相关互作网络(图3d),KEGG富集于"内吞作用"通路(图3e)。BiP-20显著下调网格蛋白介导的内吞及早期内体生物发生相关蛋白,包括EEA1、Rab5、Cltc、Vps家族、Hrs(Hgs)、Snx2、Vps35、Igf2r等,提示其诱导内体成熟阻滞与回收通路失调,可能通过破坏早期内体结构或形成杂交内体促进逃逸。
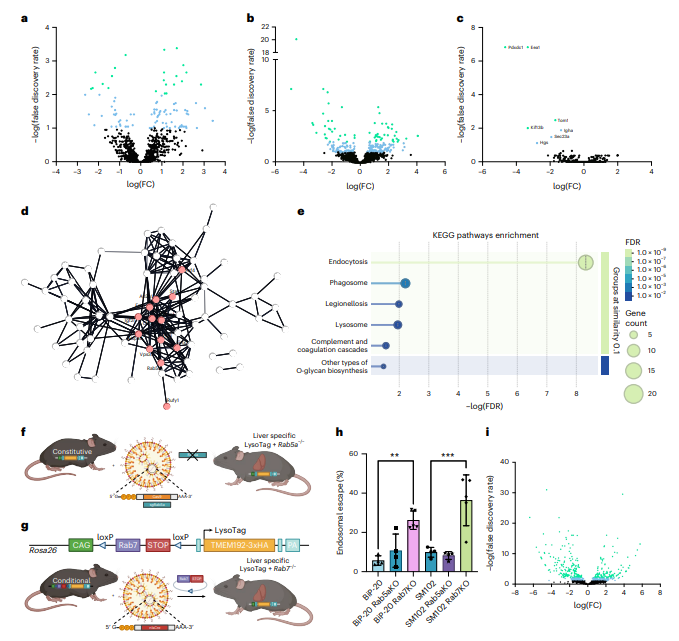
图 3. 溶酶体蛋白质组学揭示了在脂质纳米颗粒(LNP)处理后,囊泡运输和代谢过程发生了改变。
通过LNP递送sgRNA实现Rab5a敲除(67.3% indel)后,LysoBC显示逃逸效率无提升甚至略降(图3h),表明干扰早期内体形成无益于逃逸;而将LysoTag小鼠与Rab7f/f小鼠杂交并经LNP-Cre实现肝脏特异性Rab7敲除(图3g)后,BiP-20与SM102的逃逸效率分别提升约4.6倍和3.7倍(图3h),蛋白质组学证实Rab7⁻/⁻小鼠溶酶体蛋白组发生大规模重塑且降解酶减少(图3i),支持"阻断晚期内体成熟可积累逃逸能力更强的成熟缺陷型杂交内体"这一模型。