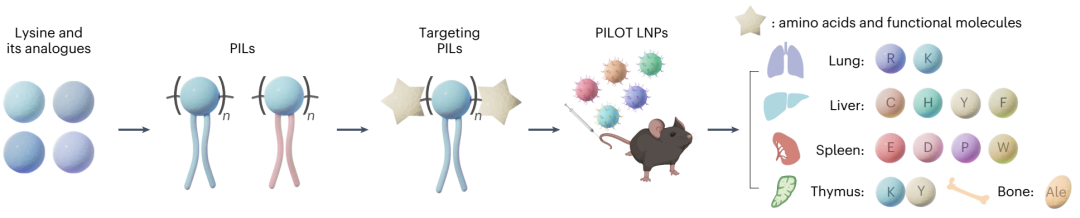
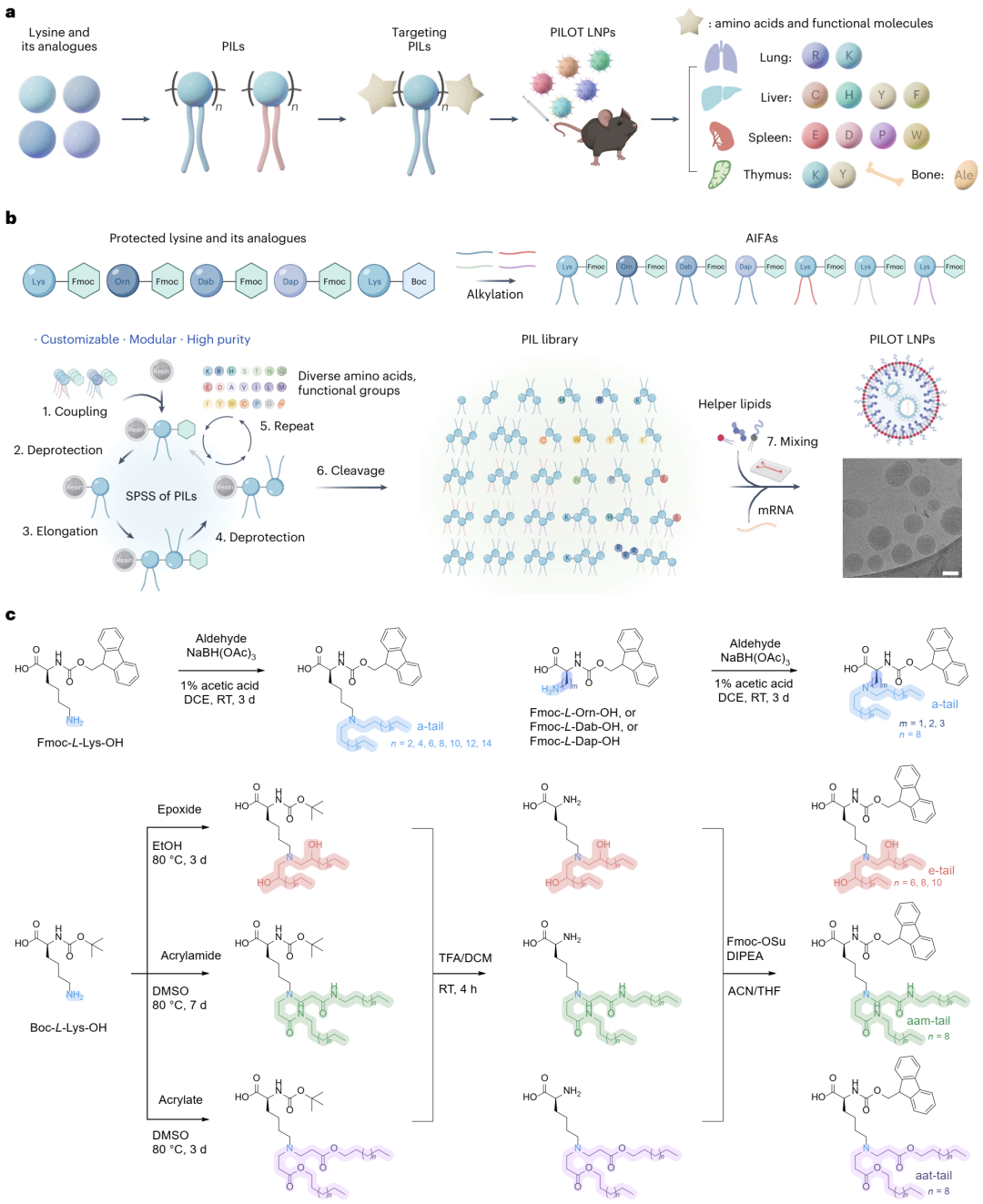
研究成功设计并合成了超过120种结构多样的PILs,这些PILs由天然氨基酸(或其它功能性构建模块)和人工烷基化离子化Fmoc保护氨基酸(AIFAs)通过SPSS方法模块化合成。具体地,通过Fmoc自脱保护反应和标准Fmoc-based SPSS技术,研究合成了包含饱和烷基链(a-tails)、含羟基烷基链(e-tails)、酰胺键(aam-tails)和酯键(aat-tails)的AIFAs。最终,研究获得了75种结构明确的PILs,命名为AnBm,其中“A”代表烷基尾链类型,“n”代表烷基链长度,“B”代表赖氨酸或其类似物,“m”代表AIFAs的数量(图1b)。
通过1H NMR和质谱验证了合成的PILs的结构,并进一步通过微流控混合技术将PILs与mRNA及辅助脂质(胆固醇、DSPC和PEG-DMG)混合制备成PIL-LNPs。cryo-TEM观察显示,a12K4 LNPs主要呈现经典的“实心核”结构,少数LNPs显示“囊泡”和“空双层囊泡”结构(图1b)。此外,TNS测定显示,随着AIFAs数量的增加,PILs的pKa值逐渐降低,这可能是由于PILs单体间通过共价酰胺键更紧密地排列,增加了离子化氨基中心间的静电排斥,从而需要更低的pH值来实现离子化(图1b)。
Fig. 1 | Construction of PIL library and PIL-based mRNA-LNPs
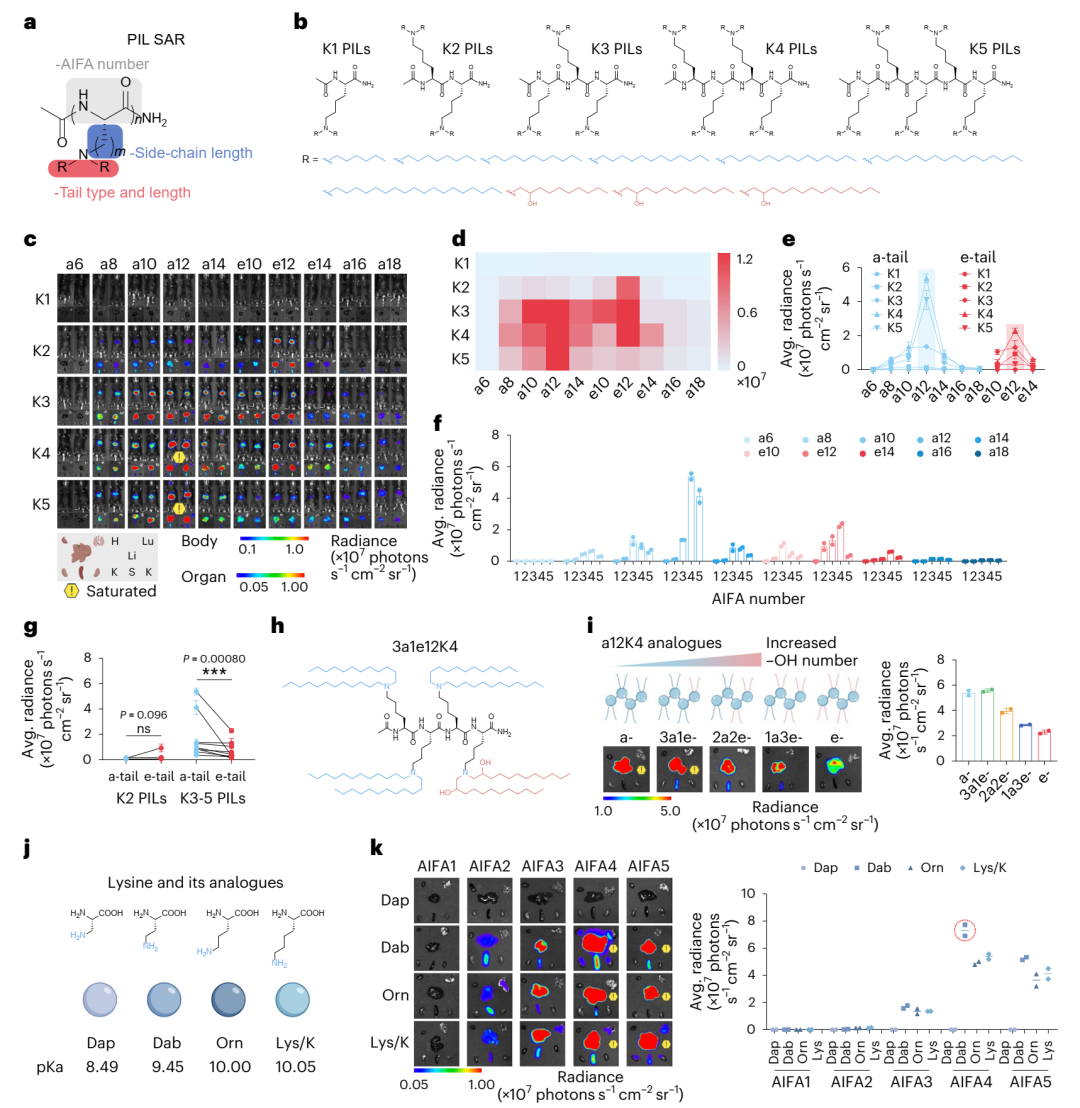
结构-活性关系(SAR),研究首先探索了PILs结构对mRNA-LNP体内递送效率的影响。通过静脉注射LNPs至C57BL/6小鼠,研究发现12碳烷基链长度的PILs表现出最高的递送效率(图2c-e)。进一步分析显示,对于a-tails和e-tails,最优的AIFA数量通常为4(图2f)。值得注意的是,a12K4在所有评估的PILs中脱颖而出,成为领先的候选结构。
为了进一步优化,研究合成了包含不同侧链长度和类型的赖氨酸类似物(如Orn、Dab和Dap)的PILs。结果显示,Dab-PILs表现出比其他类似物更优的递送效率,其中a12Dab4的Luc表达量比初始领先的a12K4高出1.36倍(图2k)。相比之下,Dap-PILs完全丧失了mRNA递送能力,突显了侧链结构和碱度对PILs设计的重要性(图2k)。
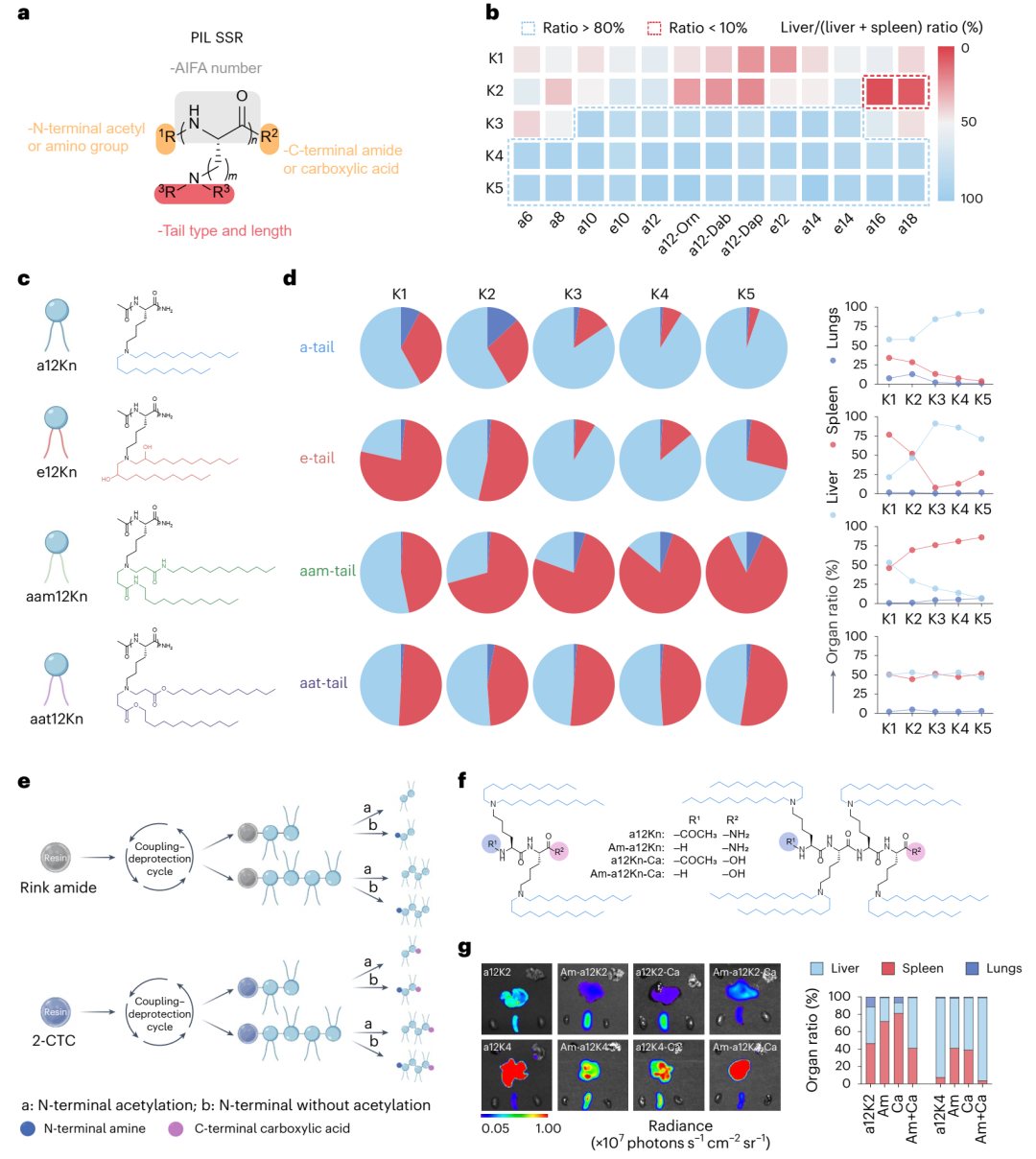
接下来,研究分析了PILs结构对组织选择性的影响。通过计算肝脏与脾脏的相对Luc表达量,研究发现含有3-4个AIFAs且烷基链长度为10-14的PILs主要介导mRNA在肝脏中的递送(>80%),这可能与其相对较低的pKa值(6.0-6.7)有关,该范围被认为对肝脏递送最优(图3b)。进一步探索不同尾链类型对组织选择性的影响,研究发现与a-tails相比,e-tails并未提高mRNA在肝脏中的递送效率,甚至在某些情况下降低了效率(图2g)。通过合成三种a12K4的混合尾链变体,研究发现增加e-tails的比例显著降低了肝脏中的mRNA表达(图2i),推测PILs中已存在的酰胺键可能消除了对额外羟基的需求。
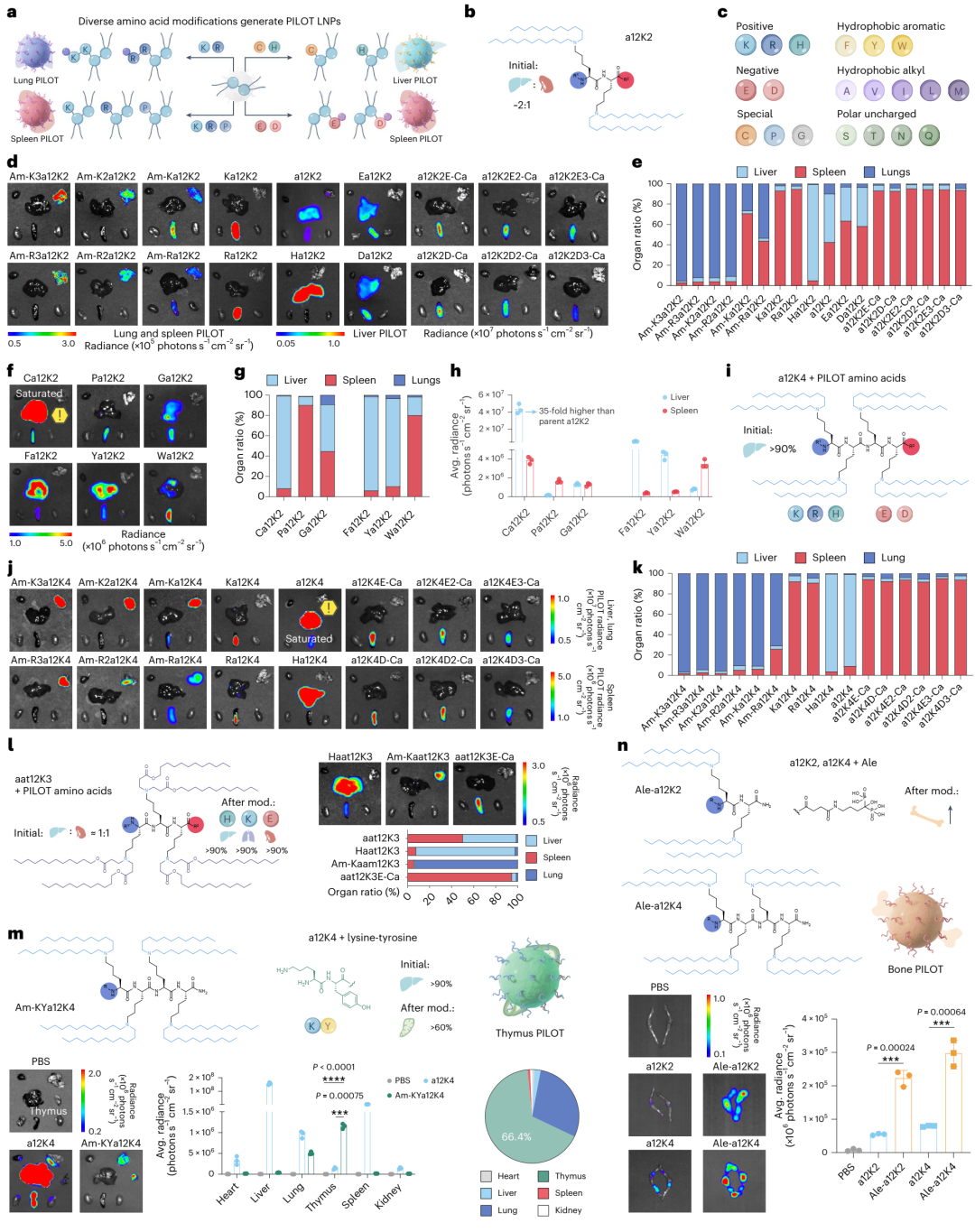
研究展示了PILOT LNPs的构建策略,即通过修饰PILs的特定氨基酸来实现对不同组织的特异性递送。研究选择了a12K2作为母体PIL,通过引入不同功能的氨基酸(如带正电的H、K、R,带负电的E、D,以及特殊氨基酸如C、P、F、Y、W等)来合成一系列PILOT LNPs(图4b, c)。例如,图4d显示了带正电氨基酸修饰的PILs(如Ha12K2, Ka12K2, Ra12K2)在肝脏和脾脏中的特异性递送,其中Ha12K2在肝脏中的特异性最高(94.5%),而Ka12K2和Ra12K2则显著提高了在脾脏中的递送效率(>93.1%)。图4e进一步验证了这些结果,通过流式细胞术分析了不同PILOT LNPs处理后小鼠肝脏、脾脏和肺中的荧光强度,证实了带正电氨基酸修饰的PILs能够显著提高在这些组织中的mRNA表达。
研究展示了特殊氨基酸和芳香族氨基酸修饰的PILs在肝脏和脾脏中的特异性递送效果。例如,Ca12K2(半胱氨酸修饰)显著提高了在肝脏中的mRNA递送效率和特异性(图4f-g),而Fa12K2(苯丙氨酸修饰)和Ya12K2(酪氨酸修饰)则分别增强了在肝脏和脾脏中的表达(图4h)。
研究还展示了多重氨基酸修饰和功能性分子引入对PILs组织选择性的影响。例如,图4m显示了通过引入赖氨酸-酪氨酸二肽(Am-KYa12K4)显著提高了在胸腺中的mRNA表达,同时减少了在其他组织中的表达。此外,图4n展示了通过引入骨靶向分子Ale(阿仑膦酸钠)显著提高了在股骨和胫骨中的mRNA表达,表明PILOT平台具有广泛的组织靶向潜力。
Fig. 4 | Development of PILOT LNPs.
PILOT LNPs在体内外递送mRNA时的稳定性和融合性对其递送效率的影响
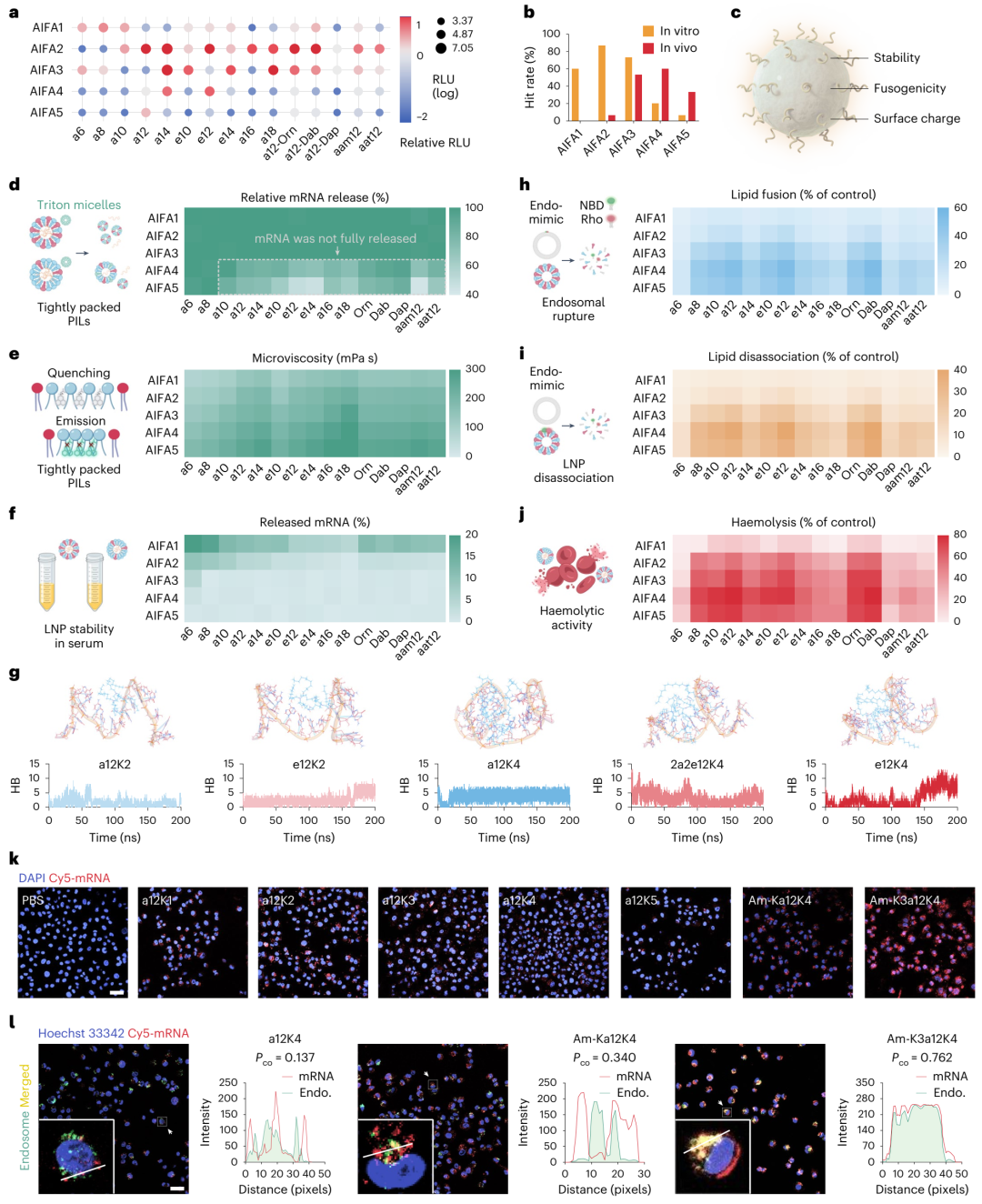
体外转染效率,与基准脂质ALC-0315相比,含有1-3个AIFAs的PILs通常表现出更高的体外转染效率(图5a, b)。然而,体内实验结果显示,含有3-5个AIFAs的PILs表现出更高的递送效率(图2f),这表明体内外性能存在显著差异。
血清稳定性实验结果显示,含有更多AIFAs和更长烷基链的PILs倾向于形成更稳定的LNPs(图5d-f),这有助于在体内循环中保持mRNA的完整性。然而,过度稳定的LNPs可能导致mRNA释放不足,从而影响递送效率(图5d)。LNPs的融合性和膜裂解活性结果显示,含有12个碳原子烷基链的PILs表现出最高的脂质融合能力(图5h, i),这有助于在体内外实现高效的mRNA递送。
融合性评估(图5h、i)表明,含有12个碳原子烷基链的PILs表现出最高的脂质融合能力。脂质融合是LNPs与细胞膜相互作用的重要过程,高融合性有助于LNPs更有效地与细胞膜融合,从而促进mRNA进入细胞。膜裂解活性评估(图5j)显示,a12K4和a12Dab4在酸性条件下(如内吞体pH 5.5)表现出最强的膜裂解活性。在细胞内吞过程中,LNPs会被包裹在内吞体中,而内吞体的酸性环境可以激活LNPs的膜裂解活性,使其破坏内吞体膜,将mRNA释放到细胞质中(图5k、l)。
综合上述结果,研究揭示了mRNA转染效率在体内外存在的差异,具体如下:体外应用的脂质纳米颗粒(LNPs)需要具备正表面电荷以实现细胞内化,具备高融合性以实现内吞体逃逸,同时具备中低稳定性以确保mRNA充分释放。因此,满足上述标准的含有1 - 2个人工合成的烷基化离子化Fmoc保护氨基酸(AIFAs)的聚离子液体(PILs),在体外表现出更优的性能。相比之下,体内给药的LNPs则需要具备高稳定性以维持血液循环,需要不同的电荷特性以形成不同的蛋白质冠组成并实现器官靶向性,同时还需要具备高融合性以实现内吞体逃逸。因此,像a12K4和a12Dab4这样更稳定且更具融合性的PILs,在体内表现出更强的mRNA递送能力。总之,这些重要的LNP特性对体内外不同过程中的各个环节有着不同程度的影响,共同决定了mRNA - LNP转染在体内外的差异。
Fig. 5 | In vitro and in vivo discrepancies of mRNA delivery by PILOT LNPs.
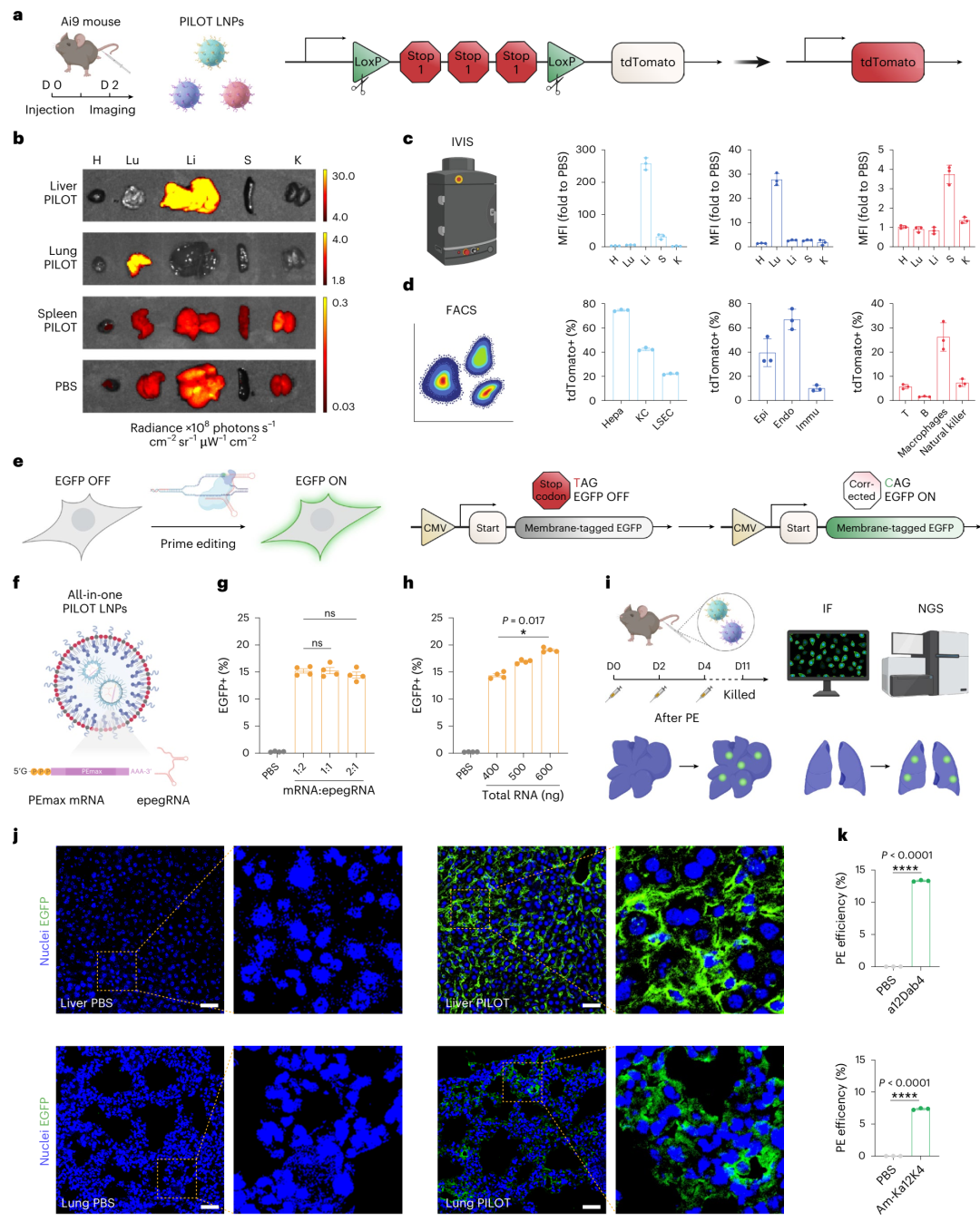
利用PILOT LNPs递送Cre重组酶mRNA,研究在Ai9报告小鼠中实现了对肝脏、肺和脾脏等组织的特异性基因编辑。生物发光成像显示,a12Dab4 Liver PILOT LNPs、Am-Ka12K4 Lung PILOT LNPs和a12K4E-Ca Spleen PILOT LNPs分别在其目标组织中实现了显著的tdTomato蛋白表达,而在其他组织中的表达可忽略不计(图6b-c)。流式细胞术分析进一步揭示了各组织中被成功编辑的细胞类型及其比例(图6d)。
通过共递送PEmax mRNA和epegRNA,研究首次实现了对肝脏和肺组织的特异性Prime编辑。利用HEK293T PE报告细胞系筛选出高效的epegRNA(图6e),研究制备了包含PEmax mRNA和epegRNA的all-in-one LNPs,并在体外验证了其编辑效率(图6g-h)。进一步,通过静脉注射PILOT LNPs至EGFP报告小鼠,研究在肝脏和肺组织中观察到了显著的EGFP阳性细胞(图6j和补充图90)。NGS分析显示,肝脏中的T-to-C点校正效率为13.3%,肺中为7.4%(图6k)。
Fig. 6 | Organ-specific gene editing by PILOT LNPs.